The key concepts of a neuron
Whether neurons are sensory or motor, big or small, they all have in common that their activity is both electrical and chemical. Neurons both cooperate and compete with each other in regulating the overall state of the nervous system, rather in the same way that individuals in a society cooperate and compete in decision-making processes. Chemical signals received in the dendrites from the axons that contact them are transformed into electrical signals, which add to or subtract from electrical signals from all the other synapses, thus making a decision about whether to pass on the signal elsewhere. Electrical potentials then travel down axons to synapses on the dendrites of the next neuron and the process repeats.
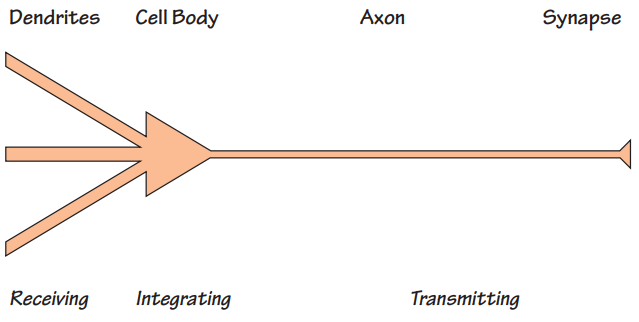
As we described in the last chapter, a neuron consists of dendrites, a cell body, an axon and synaptic terminals. This structure reflects its functional subdivision into receiving, integrating and transmitting compartments. Roughly speaking, the dendrite receives, the cell-body integrates and the axons transmit - a concept called polarization because the information they process supposedly goes in only one direction.
The key concepts of a neuron
Like any structure, it has to hold together. The outer membranes of neurons, made of fatty substances, are draped around a cytoskeleton that is built up of rods of tubular and filamentous proteins that extend out into dendrites and axons alike. The structure is a bit like a canvas stretched over the tubular skeleton of a frame tent. The different parts of a neuron are in constant motion, a process of rearrangement that reflects its own activity and that of its neighbours. The dendrites change shape, sprouting new connections and withdrawing others, and the axons grow new endings as the neuron struggles to talk a bit more loudly, or a bit more softly, to others.

3 different types of Neurons
Inside neurons are many inner compartments. These consist of proteins, mostly manufactured in the cell body, that are transported along the cytoskeleton. Tiny protuberances that stick out from the dendrites called dendritic spines. These are where incoming axons make most of their connections. Proteins transported to the spines are important for creating and maintaining neuronal connectivity. These proteins are constantly turning over, being replaced by new ones when they’ve done their job. All this activity needs fuel and there are energy factories (mitochondria) inside the cell that keep it all working. The end-points of the axons also respond to molecules called growth factors. These factors are taken up inside and then transported to the cell body where they influence the expression of neuronal genes and hence the manufacture of new proteins. These enable the neuron to grow longer dendrites or make yet other dynamic changes to its shape or function. Information, nutrients and messengers flow to and from the cell body all the time.

Dendritic spines are the tiny orange protuberances sticking out from the orange dendrites of a neuron. This is where synapses are located.
When an action potential starts at the cell body, the first channels to
open are Na+ channels. A pulse of sodium ions flashes into the cell and a
new equilibrium is established within a millisecond. In a trice, the
transmembrane voltage switches by about 100 mV. It flips from an inside
membrane voltage that is negative (about -70 mV) to one that is positive
(about +30 mV). This switch opens K+ channels, triggering a pulse of
potassium ions to flow out of the cell, almost as rapidly as the Na+ ions
that flowed inwards, and this in turn causes the membrane potential to
swing back again to its original negative value on the inside. The
action-potential is over within less time than it takes to flick a
domestic light switch on and immediately off again. Remarkably few ions
traverse the cell membrane to do this, and the concentrations of Na+ and
K+ ions within the cytoplasm do not change significantly during an action
potential. However, in the long run, these ions are kept in balance by
ion pumps whose job is to bale out excess sodium ions. This
happens in much the same way that a small leak in the hull of a sailing
boat can be coped with by baling out water with a bucket, without
impairing the overall ability of the hull to withstand the pressure of the
water upon which the boat floats.
The action potential is an electrical event, albeit a complex one. Nerve
fibres behave like electrical conductors (although they are much less
efficient than insulated wires), and so an action potential generated at
one point creates another gradient of voltage between the active and
resting membranes adjacent to it. In this way, the action potential is
actively propelled in a wave of depolarisation that spreads from one end
of the nerve fibre to the other.
An analogy that might help you think about the conduction of action
potentials is the movement of energy along a firework sparkler after it is
lit at one end. The first ignition triggers very rapid local sparks of
activity (equivalent to the ions flowing in and out of the axon at the
location of the action potential), but the overall progression of the
sparkling wave spreads much more slowly. The marvellous feature of nerve
fibres is that after a very brief period of silence (
the refractory period) the spent membrane recovers its
explosive capability, readying the axon membrane for the next action
potential.
Much of this has been known for 50 years based on wonderful experiments
conducted using the very large neurons and their axons that exist in
certain sea-creatures. The large size of these axons enabled scientists to
place tiny electrodes inside to measure the changing electrical voltages.
Nowadays, a modern electrical recording technique called
patch-clamping is enabling neuroscientists to study the
movement of ions through individual ion-channels in all sorts of neurons,
and so make very accurate measurements of these currents in brains much
more like our own.
In many axons, action-potentials move along reasonably well, but not very fast. In others, action potentials really do skip along the nerve. This happens because long stretches of the axon are wrapped around with a fatty, insulating blanket, made out of the stretched out glial cell membranea myelin sheath.

The nerve fibres above (the purple shows the axons) are wrapped in Schwann cells (red) that insulate the electrical transmission of the nerve from its surroundings. The colours are fluorescing chemicals showing a newly discovered protein complex. Disruption of this protein complex causes an inherited disease that leads to muscle-wasting.
New research is telling us about the proteins that make up this myelin
sheath. This blanket prevents the ionic currents from leaking out in the
wrong place but, every so often the glial cells helpfully leave a little
gap. Here the axon concentrates its Na+ and K+ ion channels. These
clusters of ion channels function as amplifiers that boost and maintain
the action potential as it literally skips along the nerve. This can be
very fast. In fact, in myelinated neurons, action-potentials can race
along at 100 metres per second!
Action potentials have the distinctive characteristic of being
all-or-nothing: they don’t vary in size, only in how often
they occur. Thus, the only way that the strength or duration of a stimulus
can be encoded in a single cell is by variation of the frequency of action
potentials. The most efficient axons can conduct action potentials at
frequencies up to 1000 times per second.

Alan Hodgkin and Andrew Huxley
won the Nobel Prize
for discovering the mechanism of
transmission of the nerve impulse.
They used the "giant axon"
of the squid in studies at
the Plymouth Marine Biology Laboratory